Location: Home >> Detail
J Sustain Res. 2025;7(2):e250035. https://doi.org/10.20900/jsr20250035
 ,
Gouda A. Gouda 2 ,
Tarek A. Morsy 2 ,
Hossam H. Azzaz 2 ,
Md Atikur Rahman 3 ,
Einar Vargas-Bello-Pérez 4,*
,
Gouda A. Gouda 2 ,
Tarek A. Morsy 2 ,
Hossam H. Azzaz 2 ,
Md Atikur Rahman 3 ,
Einar Vargas-Bello-Pérez 4,*
1 Department of Animal Sciences, North Carolina Agricultural and Technical State University, Greensboro, NC 27411, USA
2 Dairy Science Department, National Research Centre, 33 Bohouth St. Dokki, Giza 12622, Egypt.
3 Department of Agriculture, Nutrition and Food Systems, University of New Hampshire, Durham, NH 03824, USA
4 Facultad de Zootecnia y Ecología, Universidad Autónoma de Chihuahua, Periférico R. Aldama Km 1, Chihuahua 31031, México
* Correspondence: Ahmed E. Kholif, Einar Vargas-Bello-Pérez.
This study investigated the potential of radish seeds as a sustainable feed additive to enhance nutrient utilization and mitigate greenhouse gas emissions in ruminant systems. Five dietary treatments were formulated by supplementing radish seeds at 0, 0.5, 1.0, 1.5, and 2.0% of dietary dry matter (DM) into a basal diet consisting of concentrate feed mixture, berseem hay, and rice straw (5:4:1 ratio). Key parameters evaluated included in vitro nutrient degradability, ruminal fermentation kinetics, gas production (GP), and emissions of methane (CH4) and carbon dioxide. Gas chromatography analysis identified eucalyptol, α-pinene, and β-pinene as the predominant bioactive compounds in radish seeds. Inclusion of radish seeds led to significant linear (p < 0.001) and quadratic (p < 0.001) increases in asymptotic GP, with the highest values observed at 1.5% and 2.0% inclusion levels. Although CH4 production increased with seed supplementation, its rate exhibited a quadratic trend (p = 0.03), reaching a minimum at 1% inclusion. Radish seeds also linearly improved (p < 0.001) the degradability of DM, neutral detergent fiber, and acid detergent fiber. While total short-chain fatty acids (SCFA) and acetate concentrations increased (p = 0.004), propionate and butyrate remained unaffected. Additionally, ammonia-N concentration, microbial protein synthesis, and partitioning factor showed both linear and quadratic responses (p < 0.05 and p < 0.019, respectively). These findings highlight the potential of radish seeds to improve fermentation efficiency and reduce the environmental footprint of ruminant production systems, contributing to more sustainable livestock feeding strategies.
Animal agriculture significantly impacts the environment, particularly through greenhouse gas emissions, where methane (CH4) has a higher global warming potential than carbon dioxide (CO2) [1]. One widely explored strategy to reduce CH4 emissions is the inclusion of potential feed additives. Among these, phytogenic feed additives stand out for their dual role in reducing CH4production and improving animal health, due to their phytochemical and bioactive compounds [1,2]. Studies have shown that certain plant leaves, seeds, extracts, and essential oils, each at specific doses, vary in their effectiveness as additives in reducing CH4 production while maintaining or enhancing nutrient digestion [3].
Despite the promising nature of these additives, their variability in composition and efficacy under different conditions requires a more targeted investigation to identify specific candidates with consistent performance. There is a growing focus on identifying alternative feed ingredients for next-generation animal feed that ensure safety and efficacy, are scalable, and enhance both animal productivity and overall farm sustainability. Phytogenic feed additives, being natural and consumer-friendly, are increasingly favored over synthetic compounds, particularly as concerns over antibiotic resistance and environmental sustainability grow [4].
Extensive research is being conducted on feeding strategies and feed additives to enhance ruminant productivity, while reducing their environmental impact by improving feed efficiency and minimizing nutrient waste [5]. In previous studies by the same researchers, the effects of various phytogenic plant leaves and seeds, such as Laurus nobilis leaves, lupin seeds, Salvia officinalis shrub, turmeric rhizomes, and fennel seeds, supplemented at different levels [0, 0.5, 1, 1.5, and 2.0% of dietary dry matter (DM)] on ruminant diets were evaluated [6–8]. These studies primarily focused on their impact on nutrient digestibility, rumen fermentation, and microbial activity.
Building on this foundation, the present study investigates a less-explored candidate: radish seeds. Although radish (Raphanus sativus) is traditionally used as forage, its seeds are less commonly studied despite having a unique profile of bioactive compounds. This experiment examines whether the distinct phytochemical composition of radish seeds—including glucosinolates and isothiocyanates—can influence microbial protein synthesis, gas production (GP), and ruminal fermentation parameters compared to other phytogenic additives. The study seeks to uncover whether radish seeds can serve not only as a protein source but also as a functional additive capable of modifying rumen microbial ecology.
Radish and related vegetables like turnips (Brassica rapa) and forage kale (Brassica oleracea) are already integrated into ruminant diets in regions like New Zealand and Chile during winter grazing, when traditional forages are limited [9]. Their adaptability and dual use as both cover crops and feed sources make them ideal for sustainable ruminant production systems.
In recent years, radish seeds have gained interest as a supplementary feed ingredient in cattle diets, primarily due to their rich nutritional profile and beneficial bioactive compounds. Their high crude protein (CP; 20–25%), essential amino acids, and unsaturated fatty acids make them a potential protein-rich alternative to conventional feed sources [10]. Additionally, radish seeds contain glucosinolates, which upon hydrolysis produce antimicrobial isothiocyanates [11], potentially improving gut health. However, their anti-nutritional factors, particularly glucosinolate levels, necessitate careful dose-dependent assessment, as excessive intake can lead to adverse effects such as thyroid dysfunction and reduced feed efficiency [12]. Thus, radish seeds represent a functional feed ingredient with potential benefits and risks that must be balanced through empirical evaluation.
To date, limited studies have examined the potential of radish as an animal feed ingredient [11,13]. Therefore, this study assessed the inclusion of dried radish seeds of uniform size and weight at varying levels in a total mixed ration (TMR) on in vitro GP, CH4, and CO2 productions, and ruminal fermentation. The study hypothesis was that the secondary metabolites present in the dried radish seeds would influence ruminal microbes, modify fermentation patterns, and improve nutrient degradability.
This study was exempt from ethical review and approval by the Department of Agriculture, New Valley University, Egypt, as it was conducted entirely in vitro using ruminal fluid sourced from a local slaughterhouse. The animals were slaughtered following animal welfare rules, ensuring they experienced no pain, suffering, discomfort, or abuse.
A basal TMR was formulated using a DM basis ratio of 5:4:1 for concentrate mixture, berseem hay, and rice straw, respectively. The chemical composition of both radish seeds and TMR is presented in Table 1.
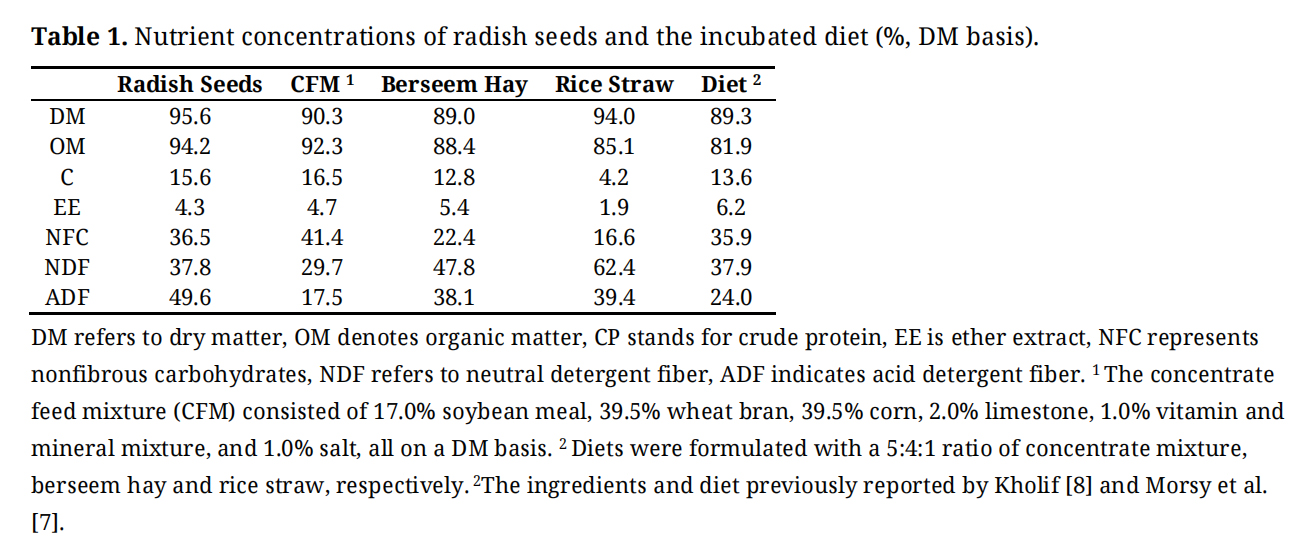 Table 1. Nutrient concentrations of radish seeds and the incubated diet (%, DM basis).
Table 1. Nutrient concentrations of radish seeds and the incubated diet (%, DM basis).
Clean, dry radish seeds (Raphanus sativus-Red) with uniform sizes and weights were ground and homogenized prior to use. To characterize the bioactive compounds relevant for dietary supplementation, the seed extract was analyzed using a Perkin Elmer Auto System XL GC/MS (J & W Agilent Technologies Inc., Palo Alto, CA, USA). However, we recommend the use of LC/MS in future work, as it is more suitable for detecting non-volatile phytochemicals such as glucosinolates and flavonoids. The analysis was performed at the Chromatography Laboratory, Central Laboratories Network, National Research Centre, Egypt, following protocols previously detailed by Kholif [8] and Morsy et al. [7].
In Vitro Fermentation and BiodegradationThe procedures for fermentation and biodegradation followed the methods described by Kholif [8] and Morsy et al. [7]. The fermentation medium was prepared according to Goering and Van Soest [14]. Sodium sulfide (2 mL) was added as a reducing agent to the buffer prior to introducing the rumen fluid. Each incubation module flask received 20 mL of ruminal inoculum along with 80 mL of the buffer solution.
Ruminal inoculum was collected from the rumen of Barki sheep (3 rams) at a local slaughterhouse in Cairo, Egypt, which were fed the same diet used as the substrate for the incubation. All supplementation levels were tested across 2 separate incubation runs, with three replicates per level. Additionally, 2 blank bottles were included in each run to determine baseline GP.
Approximately 1 g ± 10 mg of the TMR was placed in ANKOM filter bags and incubated in 250 mL ANKOM bottles equipped with a wireless GP module. Radish seeds were added at 0% (control), 0.5%, 1%, 1.5%, and 2% of the diet. Gas pressure was continuously monitored at 10-min intervals over a 48-h period. The values were then converted into gas volume (mL) under standard temperature and pressure conditions. At specific time points (2, 4, 6, 8, 10, 12, 24, 36, and 48 h), 5 mL gas samples were extracted to analyze CH4 and CO2 concentrations using a Gas-Pro detector.
The procedures for sampling and analyzing fermentation variables were conducted as outlined by Kholif [8] and Morsy et al. [7]. At 48 h of incubation, fermentation was halted by placing the bottles on ice for 5 minutes, and the pH was immediately measured. The bags were rinsed thoroughly and dried in a forced-air oven at 55 °C for 48 h. The extent of DM, NDF, and ADF degradation was calculated by comparing the post-incubation dried residue weight to the original dry weight of the substrate. GP values—including total, CH4 and CO2—were normalized to the amounts of degraded DM (dDM), NDF (dNDF), and ADF (dADF). Additionally, 5 mL of the supernatant from each bottle was collected for ammonia nitrogen (NH3-N) and short-chain fatty acid (SCFA) analysis.
Chemical AnalysisRadish seeds, individual feed ingredients, and TMR samples were analyzed for their nutrient compositions using the standard methods described by AOAC [15] and Van Soest et al. [16] as previously detailed by Kholif [8] and Morsy et al. [7].
Calculations and Statistical AnalysisThe kinetics of total GP, CH4, and CO2 were estimated by fitting the GP, CH4, and CO2 data (mL/g DM) to the nonlinear model proposed by France et al. [17] as previously detailed by Kholif [8] and Morsy et al. [7].
The partitioning factor at 48 h of incubation (PF48; mg degraded DM/mL gas) [18], and gas yield at 24 h (GY24; mL/200 mg DM) were determined. Additionally, metabolizable energy (ME) content [19], and microbial crude protein (MCP) synthesis [18] were estimated.
The statistical analysis was performed using the MIXED procedure of SAS (Version 9.4, SAS Inst., Inc., Cary, NC, USA) following a randomized block design, where each incubation run was treated as a separate block. The additive level within each block served as the experimental unit. The statistical model applied was: Yijk = μ + Li + Rj + (L × R)ij + εijk, where Yijk represents the observed value, μ is the overall mean, Li denotes the effects of radish seeds level, Rj is the run (block) effect, (L × R)ij represents the interaction between run and radish level, and εijk is the residual error. Linear and quadratic contrasts were applied to evaluate the response to increasing radish seed inclusion. Since the effects of run and its interaction with additive level were not significant (p > 0.05) for most variables, only the main effects of the additives are presented.
Proximate analysis of radish seeds and diet and volatile compounds identified in radish seeds are shown in Tables 1 and 2, respectively. Radish seeds had 15.6% CP, 37.8% NDF, and 49.6% ADF. The major volatile compounds found (% of total concentration) in radish seeds were eucalyptol (81.8%), α-Pinene (6.34%), β-Pinene (5.58%), and Fenchone (4.49%). Some other volatile compounds such as camphor, α-terpinyl acetate, and trans-caryophyllene were also observed with low concentrations (<1.0%).
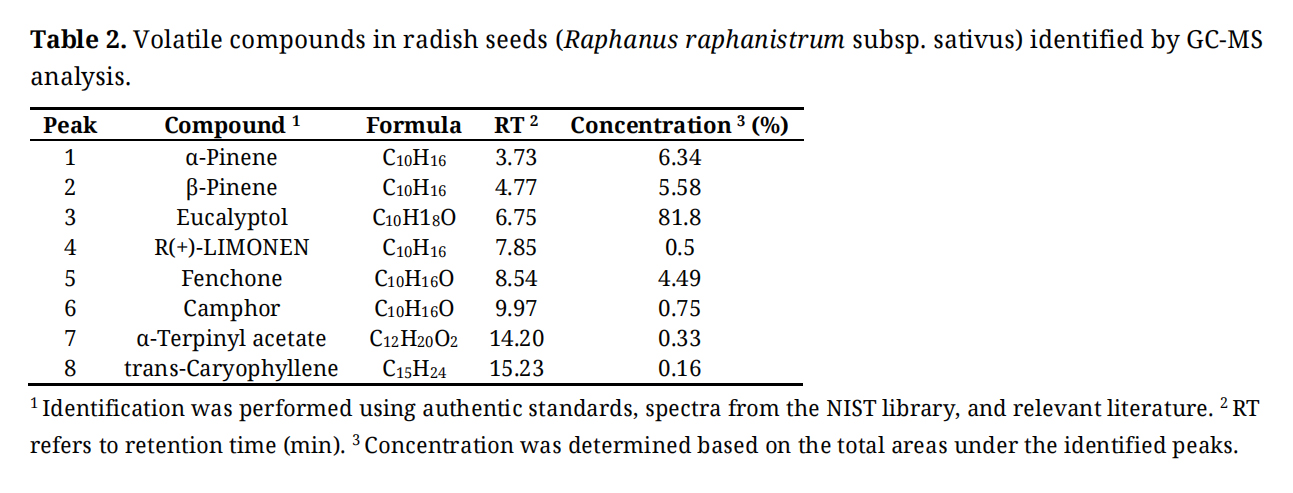 Table 2. Volatile compounds in radish seeds (Raphanus raphanistrum subsp. sativus) identified by GC-MS analysis.
Table 2. Volatile compounds in radish seeds (Raphanus raphanistrum subsp. sativus) identified by GC-MS analysis.
Figure 1 shows GP (mL/g) expressed by g of DM, dDM, dNDF, and dADF. The addition of 1.0% of radish seeds (% of DM) resulted in greater in vitro ruminal GP per g of DM, dDM, NDF, and ADF, CH4 production increased with 1.5% inclusion of radish seeds per g of DM, and then 0.5% addition of radish seeds yielded higher CH4 production per g of dDM, dNDF and dADF up to 48 h of incubation. Asymptotic GP exhibited both linear and quadratic increases as the dietary inclusion level of radish seed increased. Additionally, the rate of GP increased linearly (p = 0.002). However, no treatment effects (p = 0.14) were observed for the lag of GP.
Figure 2 shows CH4 production (mL) per g DM, dDM, dNDF, and dADF. Table 3 shows the in vitro GP, CH4, and CO2 kinetics. Asymptotic CH4 production exhibited significant linear and quadratic effects (p < 0.01) where it increased with up to 1.5% of inclusion of radish seeds and then reached the plateau at 2.0%. The rate of CH4 production (/h) had quadratic (p = 0.03) effects where it increased initially from 0 to 0.5% inclusion and then decreased afterward. The lag of CH4 production was observed to have a linear effect (p = 0.034) with the greatest GP recorded at a 1.5% inclusion level. There was no treatment effect on the proportion of CH4 production at the end of incubation (48 h).
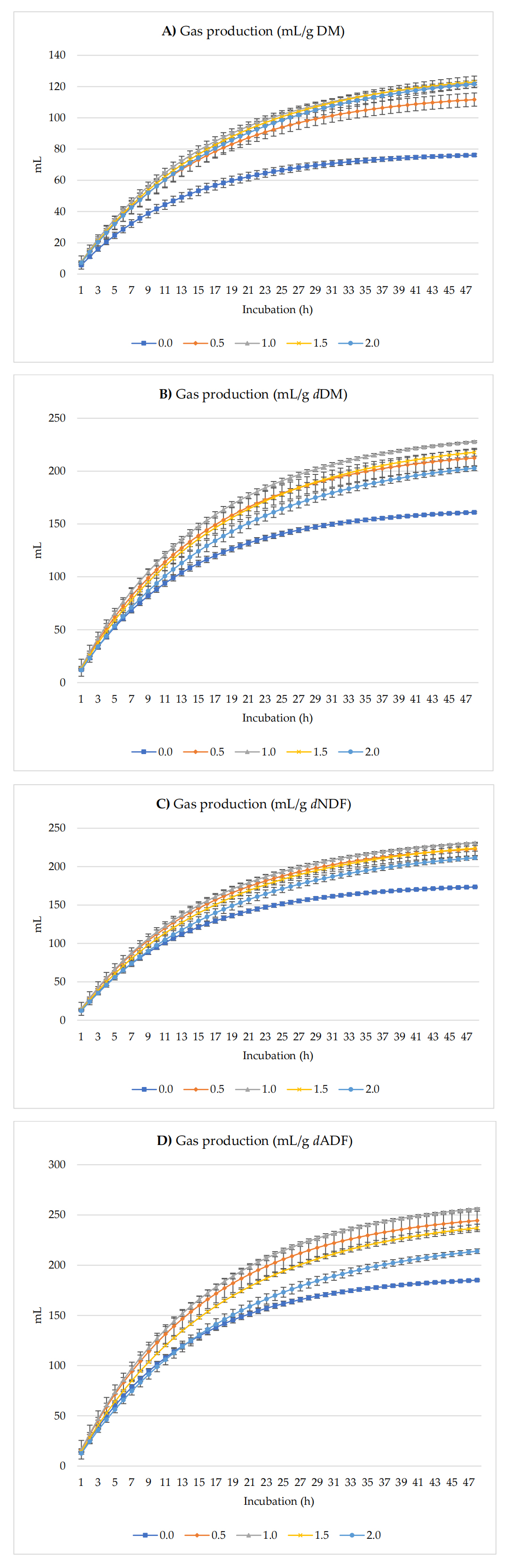 Figure 1. In Vitro ruminal GP: mL/g incubated DM (A), mL/g dDM (B), mL/g dNDF (C), and mL/g dADF (D) of a TMR supplemented with different levels of radish seeds. dDM is degradable DM, dNDF is degradable NDF, and dADF is degradable ADF.
Figure 1. In Vitro ruminal GP: mL/g incubated DM (A), mL/g dDM (B), mL/g dNDF (C), and mL/g dADF (D) of a TMR supplemented with different levels of radish seeds. dDM is degradable DM, dNDF is degradable NDF, and dADF is degradable ADF.
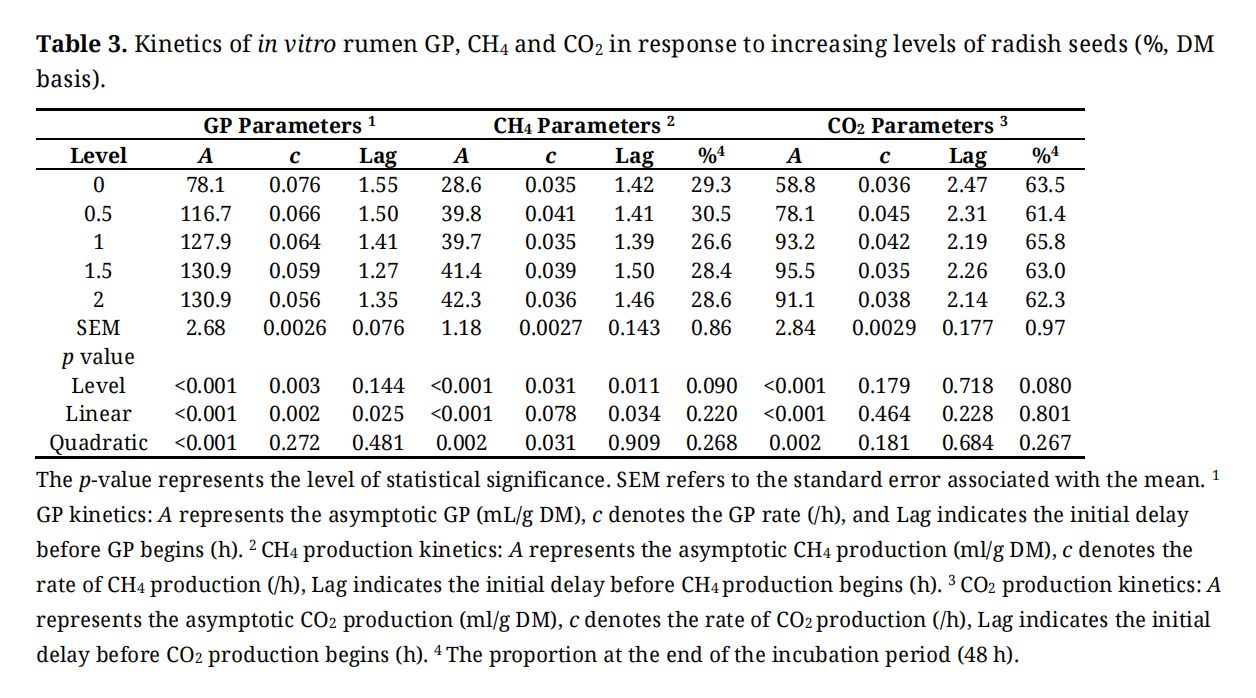 Table 3. Kinetics of in vitro rumen GP, CH4 and CO2 in response to increasing levels of radish seeds (%, DM basis).
Table 3. Kinetics of in vitro rumen GP, CH4 and CO2 in response to increasing levels of radish seeds (%, DM basis).
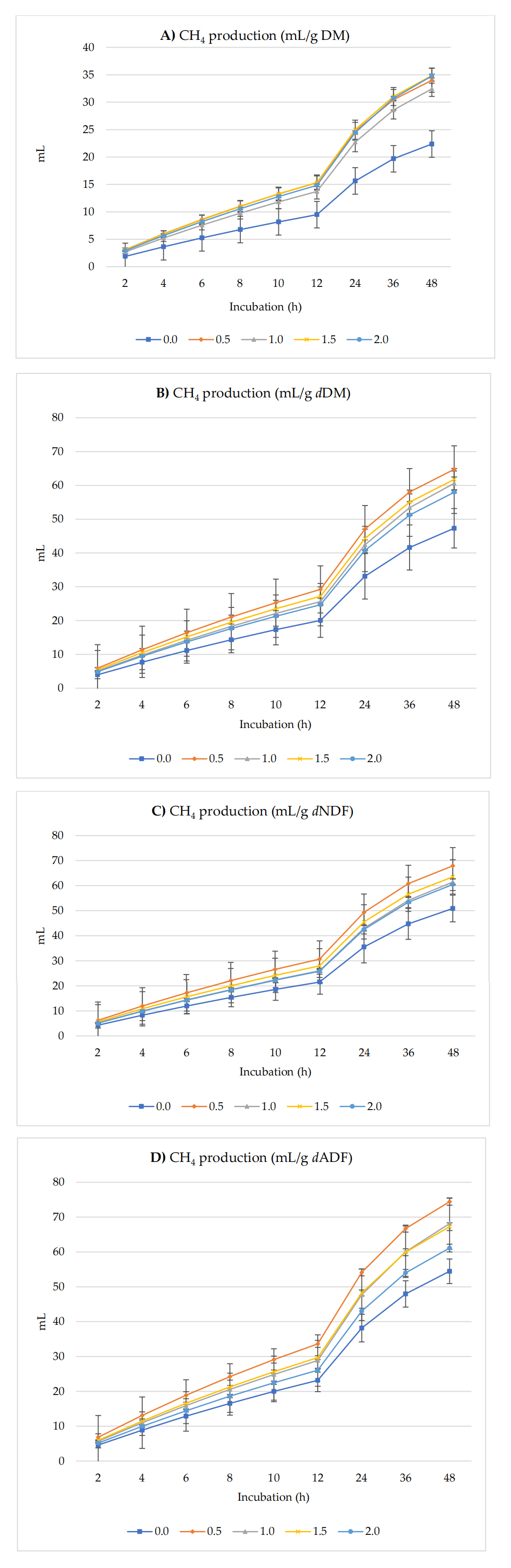 Figure 2. In Vitro ruminal methane (CH4) production: mL/g incubated DM (A), mL/g dDM (B), mL/g dNDF (C), and mL/g dADF (D) of a TMR supplemented with different levels of radish seeds. dDM is degradable DM, dNDF is degradable NDF, and dADF is degradable ADF.
Figure 2. In Vitro ruminal methane (CH4) production: mL/g incubated DM (A), mL/g dDM (B), mL/g dNDF (C), and mL/g dADF (D) of a TMR supplemented with different levels of radish seeds. dDM is degradable DM, dNDF is degradable NDF, and dADF is degradable ADF.
Figure 3 shows CO2 (mL) per g DM, dDM, dNDF, and dADF. In vitro, ruminal CO2 production increased with the supplementation of 1.0% radish seeds per g of DM, dDM, dNDF, and dADF in the diet up to 48 h of incubation. Asymptotic production of CO2 increased (p < 0.001) linearly and quadratically (p = 0.002). No effects were observed on the rate of GP (p = 0.179), lag of CO2 production (p = 0.718), and the proportion of CO2 production (p = 0.080) at the end of incubation (48 h).
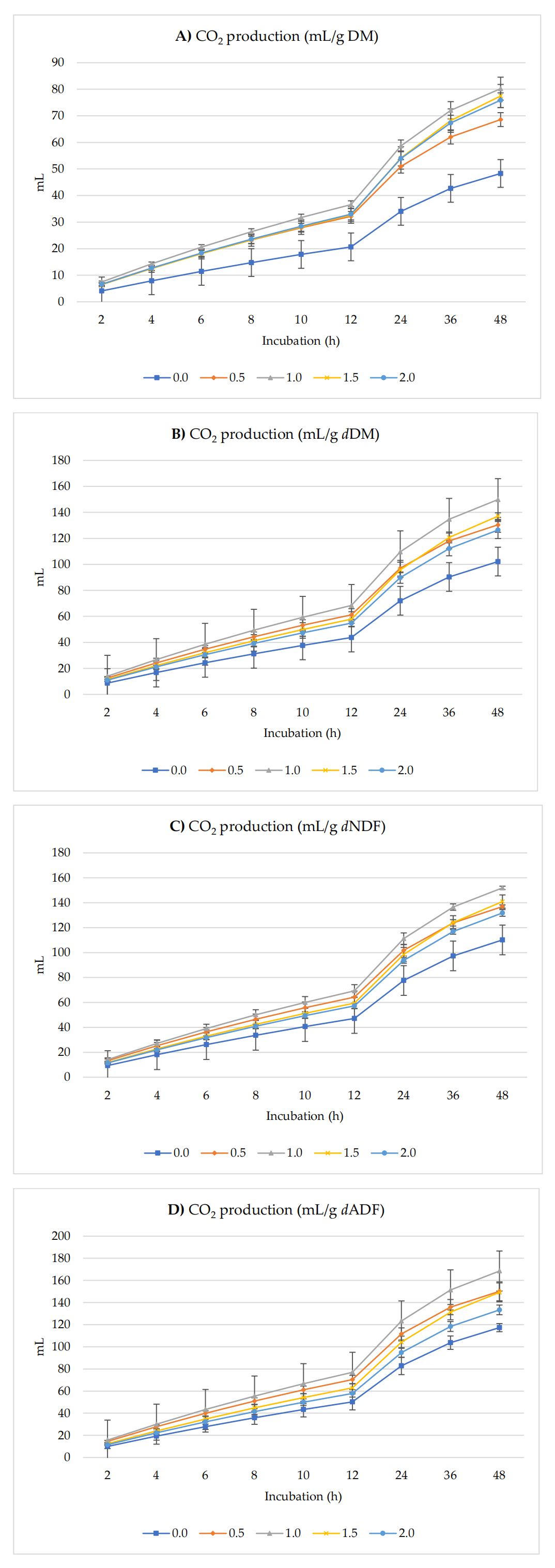 Figure 3. In Vitro ruminal carbon dioxide (CO2) production: mL/g incubated DM (A), mL/g dDM (B), mL/g dNDF (C), and mL/g dADF (D) of a TMR supplemented with different levels of radish seeds. dDM is degradable DM, dNDF is degradable NDF, and dADF is degradable ADF.
Figure 3. In Vitro ruminal carbon dioxide (CO2) production: mL/g incubated DM (A), mL/g dDM (B), mL/g dNDF (C), and mL/g dADF (D) of a TMR supplemented with different levels of radish seeds. dDM is degradable DM, dNDF is degradable NDF, and dADF is degradable ADF.
Degradability of nutrients and rumen fermentation profile are shown in Table 4. The inclusion of radish seeds linearly (p < 0.001) increased the degradability of DM, NDF, and ADF (Table 4). Whereas total SCFA and acetate increased linearly (p = 0.004), the concentration of propionate and butyrate were similar between treatments. The concentration of NH3-N, PF48, and MCP changed linearly (p < 0.01) and quadratically (p < 0.01) where it decreased initially and then increased afterward. Metabolizable energy and GY24 also changed linearly (p < 0.01) and quadratically (p < 0.01), however, it improved with the increasing amounts of radish seeds in the diets and then decreased.
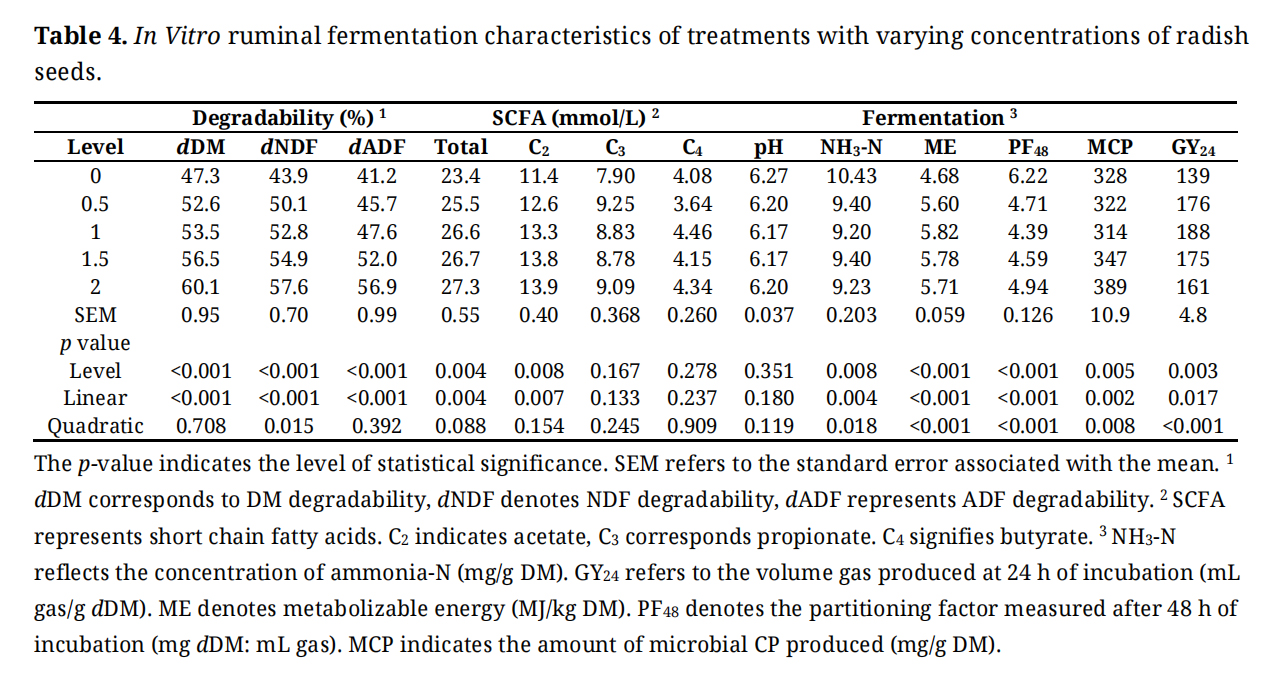 Table 4. In Vitro ruminal fermentation characteristics of treatments with varying concentrations of radish seeds.
Table 4. In Vitro ruminal fermentation characteristics of treatments with varying concentrations of radish seeds.
Radish (Raphanus sativus) is a globally used cruciferous vegetable, rich in nutrients and phytochemicals beneficial to human and animal health. It has been previously reported that the CP content of radish oil seeds can be up to 37%, with a balanced amino acid profile [20]. In our study, eucalyptol (known as 1,8-cineole) was identified as a major volatile compound, constituting 81.8% of the total volatiles. This monoterpenoid is known for its antimicrobial properties, which can influence rumen microbial populations [21].
Phytochemical concentrations in plants can vary based on factors such as geographical location, growth stage, and extraction methods [22]. Our findings align with previous reports identifying glucosinolates, phenolic compounds, and carotenoids in radish seeds [23]. These secondary metabolites contribute to plant defense mechanisms and have been associated with antioxidant, antimicrobial, anti-inflammatory, and anticancer effects [23]. For instance, glucosinolates can break down into isothiocyanates, compounds studied for their health benefits. Phenolic compounds like sinapic acid and flavonoids contribute to antioxidant activity. Additionally, radish seeds contain essential fatty acids and phytosterols, which may help lower cholesterol [24]. Plant-derived volatile compounds, such as oxygenated terpenoids, have been found to modulate rumen fermentation by affecting microbial populations. These compounds exhibited antimicrobial properties against bacteria, protozoa, and fungi by disrupting cell membrane integrity, modulating signal transduction pathways, inhibiting enzymatic activity, and interfering with bacterial colonization [25–27].
In Vitro Gas Production and Fermentation CharacteristicsIn vitro GP serves as a reliable indicator of rumen degradability, fermentation efficiency, and microbial activity. Higher GP typically reflects increased nutrient availability for rumen microbes [8]. In our study, asymptotic GP and the rate of GP increased linearly with the incremental addition of dietary radish seeds, suggesting improved ruminal fermentation and DM degradability. These findings are consistent with previous studies where the inclusion of radish by-products enhanced total GP [13].
Essential oils derived from plants including compounds like 1,8-cineole, have been shown to influence rumen fermentation and GP through their antimicrobial activity. The effectiveness of these compounds depends on their concentration and specific composition [28,29]. Higher asymptotic GP indicates effective utilization of feed by rumen microbiota, leading to increased production of SCFA, which are critical energy sources for cattle [30]. This improved fermentation supports better nutrient absorption, energy availability, and microbial protein synthesis, essential for growth, milk production, and maintenance [31].
The observed parameters in GP rate and lag time phase were similar, reinforcing the inverse relationship typically seen between lag duration and asymptotic GP [32,33]. The linear effects observed in GP kinetics suggest that the level of radish seed inclusion and their secondary metabolites were within acceptable ranges, avoiding inhibitory effects on fermentation.
Furthermore, certain constituents of plant-derived essential oils, particularly monoterpenoids like 1,8-cineole, estragole, and α-terpinyl acetate, may serve as carbon sources for specific rumen microbes due to their hydrocarbon and alcohol structures [34]. The elevated levels of volatile compounds in radish seeds could potentially modulate ruminal fermentation and GP [35]. For instance, studies have shown that eucalyptus oil, rich in eucalyptol (1,8-cineole), can reduce total gas and CH4 emissions during in vitro fermentation without affecting DM and OM digestibility [36].
Methane and Carbon Dioxide ProductionsIt was anticipated that incorporating radish seeds and their bioactive compounds would suppress CH4 formation by enhancing substrate breakdown and optimizing rumen fermentation patterns. However, our study observed an increase in CH4 production with higher levels of radish seed inclusion. Methanogenesis in the rumen involves interactions between hydrogen-producing microorganisms and methanogens, playing a crucial role in maintaining environmental homeostasis [37]. The increase in CH4 emissions could be attributed to the readily degradable nature of radish seeds, providing additional carbon sources for methanogens [38]. Additionally, a linear increase in total SCFAs and acetate was observed without affecting propionate levels, aligning with the observed increased CH4 production. Fermentation of feedstuffs into acetate generates excess hydrogen, which methanogens utilize for CH4 production [39].
In this study, the increase in rumen CO2 can be explained as a byproduct of ruminal microbial fermentation, where microbes break down ingested feed components like radish carbohydrates, producing CO2 alongside acetate, propionate, and butyrate [40].
Degradability and Nutrient UtilizationMonitoring rumen pH is vital for assessing the equilibrium and overall health of the ruminal ecosystem. In the current study, pH levels remained stable across different radish seeds inclusion rates, ranging between 6.17 and 6.27, within the normal range of 5.0 to 7.5 [41]. Total SCFA concentration increased with radish seed supplementation, correlating with improved nutrient degradability. Higher SCFA levels indicate enhanced fermentability and better nutrient acceptability, stimulating ruminal microbial growth and activity [42,43]. Short-chain fatty acid serve as a significant of the energy source for ruminants, potentially supplying up to 80% of their daily energy needs [44]. They also demonstrate a strong correlation with ME and degradable OM [19]. Increased SCFA levels observed in the treatments suggest improved fermentability and nutrient utilization [45,46].
Specifically, acetate level increased with higher inclusion rates of inclusion of radish seeds. Acetate is crucial for cattle as it serves as a primary precursor for energy production and milk fat synthesis [47]. The microbial breakdown of fibrous feed components in the rumen leads to acetate formation, contributing to approximately 50–70% of the total SCFAs, which are major energy sources for ruminants [48]. In dairy cows, high rumen acetate levels are directly correlated with enhanced milk fat production, as acetate provides the necessary acetyl-CoA units for de novo fatty acid synthesis in the mammary gland [49]. Additionally, acetate supports gluconeogenesis and provides energy for maintenance and growth [50].
Digestible DM serves as an important indicator of a feed’s nutritive value and its potential to supply energy to livestock [51,52]. Alongside dADF and dNDF, dDM reflects the extent to which various feed fractions are decomposed by rumen microbes. The observed enhancements in dDM, dADF, and dNDF across all levels of radish seed inclusion highlight the beneficial presence of bioactive compounds in radish. These improvements may be attributed to the intrinsic properties of radish seeds, particularly their phytochemicals, which likely promoted microbial growth and activity, enhancing diet colonization and degradation [53].
Radish seeds contain bioactive phytochemicals that contribute to their potential as a functional additive in ruminant feeding programs. Active compounds in radish seeds, such as 1,8-cineole, may stimulate rumen microflora, enhancing their ability to produce essential vitamins and enzymes that optimize digestibility [34]. For instance, supplementing Bermuda grass hay with San wormwood essential oil, containing 56.7% 1,8-cineole, enhanced populations of Ruminococcus albus and Streptococcus bovis, both crucial for cellulose and hemicellulose digestion [54,55]. Importantly, the improvements in dDM, dNDF, and dADF degradability were not solely due to the radish seeds’ nutritional content but were more likely driven by their functional bioactive compounds enhancing microbial activity in the rumen. Consequently, radish seeds phytochemicals may have facilitated digestibility, with greater effects observed at higher supplementation levels [2,34]. These enhancements suggest that radish seeds can improve nutrient and energy availability for animals. Our in vitro findings align with previous reports indicating that radish supplementation increases SCFA concentration, DM degradability, and total GP [13]. These characteristics demonstrate radish’s potential to improve rumen fermentation. Further in vivo studies are recommended to validate these findings under practical farm conditions. Additionally, conducting rumen microbiome analyses would help determine the impact of radish on rumen microorganisms as rapid fermentation may lead to decreased fibrolytic bacterial populations due to a shift toward amylolytic or proteolytic dominance [56].
Ammonia-N is essential for rumen microbial function, supporting the breakdown and utilization of feed components during digestion and metabolism. As a byproduct of protein degradation, including both plant and microbial proteins, NH3-N serves as a key N source that rumen microbes utilize for microbial protein synthesis. Following microbial cell turnover or passage from the rumen, this microbial protein becomes an important source of absorbable dietary protein for ruminants [57]. The reduction in NH3-N concentrations may be attributed to the presence of 1,8-cineole, a bioactive compound known to suppress hyper-ammonia-producing bacteria and reduce ruminal NH3-N levels [36,58]. However, this reduction was not observed in the present study, possibly due to the relatively low concentration of 1,8-cineole in radish seeds or the specific chemical composition of the incubated substrates.
Nevertheless, the observed decline in NH3-N concentration in conjunction with increased in MCP production suggests that available ammonia was efficiently utilized by rumen microbes. This implies that microbial populations actively incorporated NH3-N into cellular protein structures, thereby enhancing the efficiency of nitrogen utilization [59]. The presence of ciliate protozoa typically contributes to bacterial nitrogen recycling, yet their population tends to decline when dietary protein degradation increases or when specific plant secondary metabolites are present. It is plausible that the phytochemical composition of radish seeds contributed to reduced protozoal counts and inhibited hyper-ammonia-producing bacteria [34,37], explaining the lower NH3-N concentrations and the concurrent rise in MCP production. Microbial crude protein is a crucial amino acid source for ruminants and its synthesis is influenced by multiple factors, including the synchrony of fermentable energy and nitrogen availability, rumen passage rate, and microbial recycling efficiency [60]. The inclusion of moderate levels of readily fermentable carbohydrates —such as those present in radish seeds—likely enhanced substrate availability, stimulated microbial growth in the liquid phase of rumen contents, and ultimately increased MCP synthesis and OM degradability [60,61]. These improvements indicate a better rumen environment for microbial proliferation and activity.
Furthermore, radish seeds appear to modulate the rumen microbial ecosystem by selectively inhibiting non-beneficial microorganisms while supporting beneficial bacterial growth, likely due to their rich phytochemical profile. This microbial shift enhances fermentation efficiency and nutrient digestibility. The observed increased ME, PF48, and MCP levels suggest an improved energy-nitrogen balance in the rumen, fostering more efficient microbial biomass production [7]. Moreover, certain seed-derived phytochemicals in radish seeds may influence aromatic amino acid biosynthesis pathways, further supporting enhanced microbial growth and nitrogen utilization [62]. Increased rumen ME availability observed in this study is of particular importance in ruminant production systems, as ME represents the energy fraction available to the host after losses in feces, urine, and CH4. Optimizing ME can improve growth rates, milk production, and reproductive performance [63], especially under energy-demanding physiological states such as lactation or rapid growth [64].
The increase in GY24, particularly per unit of digested DM, in diets supplemented with radish seeds supports the hypothesis that radish improves fermentative efficiency. Higher GY24 values indicate a more favorable microbial environment and greater availability of fermentable substrates [57]. This may be attributed to both the nutritional composition of radish seeds and their bioactive compounds, which potentially accelerate microbial metabolism and stimulate SCFA production. However, it is essential to interpret GP cautiously, as elevated gas output may also reflect increased CH4 formation unless paired with favorable SCFA profiles and reduced methanogenesis.
Based on the findings of this study, the null hypothesis is rejected. The data demonstrate that secondary metabolites in dried radish seeds significantly influenced ruminal microbial activity, altered fermentation patterns (as shown by increased GP, SCFA, and CH4), and enhanced nutrient degradability (dDM, dNDF, dADF). These effects confirm that radish seeds are not inert in the rumen but actively contribute to improving fermentation dynamics and nutrient utilization. The observed increases in SCFA, MCP, and ME, along with improved digestibility parameters, directly support the initial hypothesis and underscore the functional role of radish-derived phytochemicals in modulating rumen function.
Overall, this study demonstrated that incremental amounts (from 0.5 to 2% DM) of radish seeds improved in vitro ruminal fermentation, total SCFA, but increased CH4 production. Radish seeds could be a simple, sustainable solution to reducing the environmental impact of livestock farming while maintaining efficient animal production. Further studies should consider expanding our findings with in vivo studies in different production systems and perhaps analyzing the effect of radish seeds on physicochemical and sensorial characteristics of final products such as meat and milk.
Conceptualization, AEK, HHA and GAG; Data curation, AEK, TAM, EVBP, MAR and GAG; Formal analysis, AEK, HHA and TAM; Funding acquisition, AEK; Investigation, AEK, TAM and GAG; Methodology, AEK, TAM, HHA and GAG; Project administration, AEK; Resources, AEK and GAG; Software, AEK, MAR, EVBP and TAM; Supervision, AEK and TAM; Validation, AEK, EVBP and GAG; Visualization, AEK and TAM; Writing—original draft, AEK, EVBP and MAR; Writing—review and editing, AEK All authors have read and agreed to the published version of the manuscript.
This research received no external funding.
The study's original contributions are incorporated within the article, and any additional inquiries can be directed to the corresponding authors.
The authors declare no conflicts of interest.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58.
59.
60.
61.
62.
63.
64.
Kholif AE, Gouda GA, Morsy TA, Azzaz HH, Rahman MA, Vargas-Bello-Pérez E. Sustainable modulation of rumen fermentation and greenhouse gas emissions using radish seeds as a natural feed additive. J Sustain Res. 2025;7(2):e250035. https://doi.org/10.20900/jsr20250035.

Copyright © Hapres Co., Ltd. Privacy Policy | Terms and Conditions




