Location: Home >> Detail
J Sustain Res. 2025;7(4):e250064. https://doi.org/10.20900/jsr20250064
 ,
Gouda A. Gouda 2 ,
Tarek A. Morsy 2 ,
Uchenna Y. Anele 1
,
Gouda A. Gouda 2 ,
Tarek A. Morsy 2 ,
Uchenna Y. Anele 1
1 Department of Animal Sciences, North Carolina Agricultural and Technical State University, Greensboro, NC 27411, USA
2 Dairy Science Department, National Research Centre, 33 Bohouth St. Dokki, Giza 12622, Egypt
* Correspondence: Ahmed E. Kholif
This study investigated the sustainable effects of incorporating graded levels of dried cress seeds [Lepidium sativum; 0, 0.5, 1.0, 1.5, and 2.0% dry matter (DM) basis] into a total mixed ration (TMR) containing 500 g concentrate mixture, 400 g berseem hay, and 100 g rice straw on in vitro ruminal gas production (GP), methane (CH4) and carbon dioxide (CO2) kinetics, nutrient degradability, and fermentation characteristics. Gas chromatography–mass spectrometry analysis identified a complex volatile profile in cress seeds dominated by 1,8-cineole (61.8%), estragole (15.1%), fenchone (7.71%), and pinenes, compounds known for antimicrobial and rumen-modulating properties. In vitro fermentation results showed a quadratic increase with cress seeds inclusion (p < 0.01) in GP parameters by 50.2%, from 78.1 to 117.3 mL/g DM, while the GP rate decreased with cress seeds inclusion by 9.2% (p = 0.007). Methane production linearly increased by 38.1% (p = 0.006) but the CH4 production rate declined by 34.3% (p = 0.026), leading to a 20.8% linear reduction (p = 0.002) in CH4 proportion relative to total gas, indicating improved fermentation efficiency and CH4 mitigation. CO2 production parameters quadratically increased, with asymptote rising by 42.2% (p < 0.01) and rate by 50.0% (p = 0.005). Nutrient degradability was markedly enhanced with cress seed inclusion. DM degradability quadratically increased by 38.7% (p = 0.041), neutral detergent fiber (NDF) degradability by 39.9% (p < 0.001), and acid detergent fiber (ADF) degradability by 43.6% (p < 0.001). Total volatile fatty acids (VFAs) linearly increased by 29.5% (p < 0.001), with acetate and propionate concentrations rising linearly by 36.0% (p < 0.001) and 23.9% (p = 0.002), respectively, while the acetate-to-propionate ratio remained stable (p = 0.397). Metabolizable energy (ME) improved quadratically by 18.6% (p = 0.003), and microbial crude protein (MCP) production linearly increased by 36.6% (p < 0.001). Ruminal pH and ammonia nitrogen concentrations remained unaffected. As a conclusion, dietary inclusion of dried cress seeds up to 2% of the diet enhances ruminal fermentation efficiency and nutrient degradability, reduces CH4 intensity, and stimulates microbial protein synthesis, highlighting its potential as a natural feed additive to improve ruminant productivity and reduce greenhouse gas emissions.
Ruminant livestock production plays a crucial role in global food security by providing high-quality protein through meat and milk production. However, these animals are also a major source of enteric CH4, a potent greenhouse gas contributing to climate change [1]. Methane emissions not only exacerbate environmental concerns—accounting for 17% of global anthropogenic greenhouse gas emissions—but also represent an energy loss of up to 12% of gross energy intake, thereby negatively affecting animal productivity and feed efficiency [2]. Therefore, there is a pressing need to develop nutritional strategies that reduce CH4 emissions without compromising animal health or performance.
One promising approach involves the use of natural feed additives derived from plants rich in bioactive compounds such as essential oils, tannins, and saponins. These phytochemicals can selectively inhibit methanogenic archaea and protozoa, shift fermentation toward propionate (C3) production, and enhance fiber digestibility [3]. Essential oils and terpenoids, in particular, have demonstrated antimicrobial and rumen-modulating properties that can improve fermentation efficiency and reduce greenhouse gas output [4].
Lepidium sativum, commonly known as cress seed, is an herbaceous plant widely cultivated for its nutritional and medicinal benefits. The seeds contain a complex profile of volatile compounds, including oxygenated monoterpenes like 1,8-cineole, estragole, and pinene isomers, which exhibit antimicrobial, antioxidant, and anti-inflammatory activities [5]. Although these properties suggest cress potency for rumen modulation, there is limited research on how cress seed supplementation affects ruminal fermentation dynamics, CH4 and CO2 production, and nutrient degradability.
Incorporating cress seeds into ruminant diets could represent a sustainable and natural strategy to improve feed utilization and reduce CH4 emissions, aligning with the goals of environmentally responsible livestock production. Previous studies have shown that supplementing ruminant diets with Lepidium sativum seeds can enhance rumen fermentation, improve nutrient digestibility, and increase milk yield [6,7].
We hypothesized that the supplementation of dried cress seeds in the diet will positively modulate ruminal fermentation by enhancing nutrient degradability and VFAs production, while simultaneously reducing CH4 emission intensity through its bioactive phytochemical constituents. Accordingly, this study aimed to investigate the effects of incorporating varying levels of dried cress seeds into a TMR on in vitro GP, including CH4 and CO2 emissions, as well as ruminal fermentation characteristics.
A basal TMR was formulated as the substrate for the in vitro study, consisting of (per kg DM): 500 g concentrate mixture, 400 g berseem hay, and 100 g rice straw. The chemical composition of the TMR, diet ingredients, and cress (Lepidium sativum) seeds is presented in Table 1.
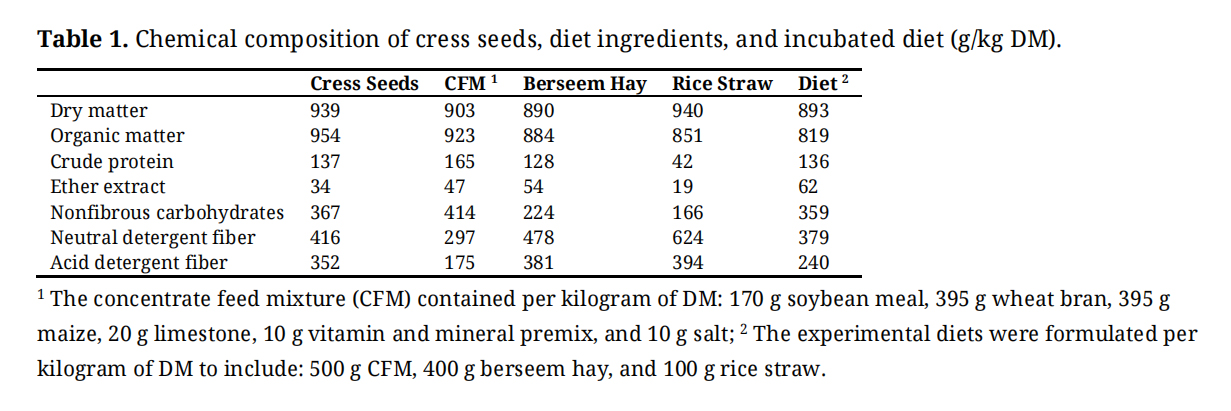 Table 1. Chemical composition of cress seeds, diet ingredients, and incubated diet (g/kg DM).
Table 1. Chemical composition of cress seeds, diet ingredients, and incubated diet (g/kg DM).
Clean, air-dried cress seeds were sourced from a local supplier in Egypt (Harraz, Cairo, Egypt). Prior to use, the seeds were ground and thoroughly homogenized. The essential oil profile of the cress seeds was analyzed at the Central Laboratory of the National Research Centre (Egypt) using a Perkin Elmer Auto System XL gas chromatograph–mass spectrometer (GC–MS) equipped with a ZB-5 capillary column (60 m × 0.32 mm i.d.; Agilent, USA). The injector temperature was initially held at 50 °C for 1 min and then increased to 240 °C at a rate of 3 °C/min. Helium served as the carrier gas at a flow rate of 1 mL/min, with a split ratio of 1:10. The column effluent was introduced directly into the mass spectrometer operated in electron ionization (EI) mode at 70 eV. Mass spectra were recorded over a range of 40–300 amu with a scan time of 1 s. Tentative identification of compounds was conducted by comparing retention times and mass spectra with entries in the NIST and WILEY mass spectral libraries.
In Vitro Fermentation and BiodegradationThe in vitro fermentation medium was prepared following the protocol described by Goering and Van Soest [8]. A reducing solution containing sodium sulfide (2 mL) was added to the buffer immediately prior to the addition of the ruminal inoculum. Each 250 mL incubation bottle received 20 mL of rumen fluid and 80 mL of the buffer solution.
Rumen fluid was collected from three sheep slaughtered at a local abattoir in Cairo, Egypt. For at least two months prior to slaughter, the animals had been fed a diet consisting of concentrate mixture, berseem hay, and rice straw at a ratio of 500:400:100 (DM basis), with ad libitum access to water. Rumen contents were sampled, stored, and processed according to the standardized procedures outlined by Fortina et al. [9]. All necessary permissions for the use of rumen fluid from slaughtered animals were obtained from the slaughterhouse. Rumen fluid was collected within 10 mins postmortem by hand-squeezing approximately 150–250 g of digesta through a colander into a beaker, repeating the process until ~1000 mL was obtained. The fluid was filtered through two layers of cheesecloth to remove large particles, and the solid residues were squeezed to extract microbe-rich fluid from feed particles. The initial pH of the inoculum ranged from 6.8 to 6.9.
Each treatment was evaluated in two incubation runs, with three replicates per treatment per run. Two blank bottles (inoculum without substrate) were included in each run to account for baseline GP. Approximately 1.00 ± 0.01 g of TMR was weighed into ANKOM F57 filter bags (Ankom Technology, Macedon, NY, USA) and placed into 250 mL ANKOM bottles connected to an automated wireless GP system (AnkomRF GP System, Ankom Technology, USA), equipped with built-in pressure sensors. Dried cress seeds were included in the TMR at 0 (control), 0.5, 1.0, 1.5, and 2.0% of diet DM. The cress seeds were used as feed additives and added in the incubation bags without replacing any component of the concentrate or forage mixture. During incubation, the bottles were continuously agitated in a shaking incubator (39 ± 0.5 °C) to simulate rumen mixing and maintain uniform fermentation conditions. Gas pressure was recorded every 10 mins over a 48-h incubation period and converted to gas volume (mL) under standard temperature and pressure conditions. Net GP was calculated by subtracting the gas volume recorded from the blanks.
At 2, 4, 6, 8, 10, 12, 24, 36, and 48 h of incubation, gas samples (5 mL) were withdrawn from the sampling ports and analyzed for CH4 and CO2 concentrations using a Gas-Pro detector (CROWCON Tetra3 Gas Analyzer, Abingdon, UK). Measurements were taken at 2-h intervals up to 12 h to capture the early fermentation phase, and then at 12-h intervals thereafter to monitor CH4 production during the later stages of incubation.
Kinetics of total GP, CH4, and CO2 (mL/g DM) were estimated by fitting the cumulative production data to the nonlinear model described by France et al. [10], using the NLIN procedure of SAS (Version 9.4; SAS Institute Inc., Cary, NC, USA). The model used was: y = A × [1 − e−c (t−Lag)] where y is the cumulative volume of GP, CH4, or CO2 (mL/g DM) at time t (h); A is the asymptotic GP (mL/g DM); c is the fractional rate of GP (/h); and Lag (h) represents the discrete lag time prior to gas release.
The partitioning factor at 48 h (PF48; mg degraded DM/mL gas) was calculated as described by Blümmel et al. [11], and gas yield at 24 h (GY24; mL/200 mg DM) was also determined. ME was estimated using the equation of Menke et al. [12], and MCP production was calculated following Blümmel et al. [11].
Sampling and Analysis of Fermentation VariablesAfter 48 h of incubation, fermentation was halted by placing the bottles on ice for 5 mins, followed by immediate pH measurement using a calibrated pH meter (Thermo Scientific, Orion StarTM A121, Beverly, MA, USA). The ANKOM F57 filter bags were then removed and dried in a forced-air oven at 55 °C for 48 h. DM, NDF, and ADF degradability were determined by calculating the difference between the initial and final weights of the dried substrates. Total gas, CH4, and CO2 production were expressed relative to the amount of degraded DM (dDM), NDF (dNDF), and ADF (dADF) after 48 h of incubation.
Fermented fluid samples (5 mL) were collected from each bottle for the determination of NH3-N and total and individual VFAs concentrations. A 3 mL subsample was mixed with 3 mL of 0.2 M hydrochloric acid for NH3-N analysis, following the method described by AOAC [13]. For VFA determination, an 0.8 mL aliquot of the fermentation fluid was combined with 0.2 mL of a metaphosphoric acid solution (250 g/L) and analyzed using steam distillation followed by titration.
Chemical AnalysisSamples of cress seeds and the TMR were analyzed for their chemical composition using standard procedures. Ash content was determined by incinerating the samples in a muffle furnace at 550 °C for 12 h (AOAC method ID 942.05). Crude protein (CP) was analyzed using the Kjeldahl method (AOAC method ID 954.01), and ether extract (EE) was determined using Soxhlet extraction with diethyl ether (AOAC method ID 920.39), as described by AOAC [13].
Neutral detergent fiber was analyzed without the use of alpha-amylase but with sodium sulfite, following the method of Van Soest et al. [14]. Acid detergent fiber was measured according to AOAC [13] (method ID 973.18) and reported exclusive of residual ash. Non-structural carbohydrates, cellulose, hemicellulose, and organic matter (OM) concentrations were subsequently calculated based on the primary analytical results.
Statistical AnalysisStatistical analysis was conducted using the MIXED procedure of SAS (Version 9.4; SAS Institute Inc., Cary, NC, USA), following a randomized block design in which each incubation run was considered a separate block. The additive level within each block was treated as the experimental unit. The statistical model used was: Yijk = μ + Li + Rj + (L × R)ij + εijk, where Yijk is the observed value, μ is the overall mean, Li represents the fixed effect of cress seed level, Rj is the random effect of run (block), (L × R)ij is the interaction between cress level and run, and εijk is the residual error. Linear and quadratic polynomial contrasts were employed to assess the response to increasing levels of cress seed inclusion. As neither the run effect nor its interaction with additive level was statistically significant (p > 0.05) for most variables, only the main effects of additive levels are reported.
The GC–MS analysis of Lepidium sativum seeds revealed a diverse profile of volatile compounds, dominated by oxygenated monoterpenes and phenylpropanoids (Table 2). The most abundant compound was 1,8-cineole, representing 61.8% of the total volatile fraction. Other notable constituents included estragole (15.1%), fenchone (7.71%), α-pinene (6.96%), and β-pinene (6.72%). Minor compounds such as limonene (1.06%), camphor (0.22%), 2-phenylethyl-1,1,2,2-d4-amine (0.28%), α-terpinyl acetate (0.07%), and trans-caryophyllene (0.08%) were also detected in lower concentrations.
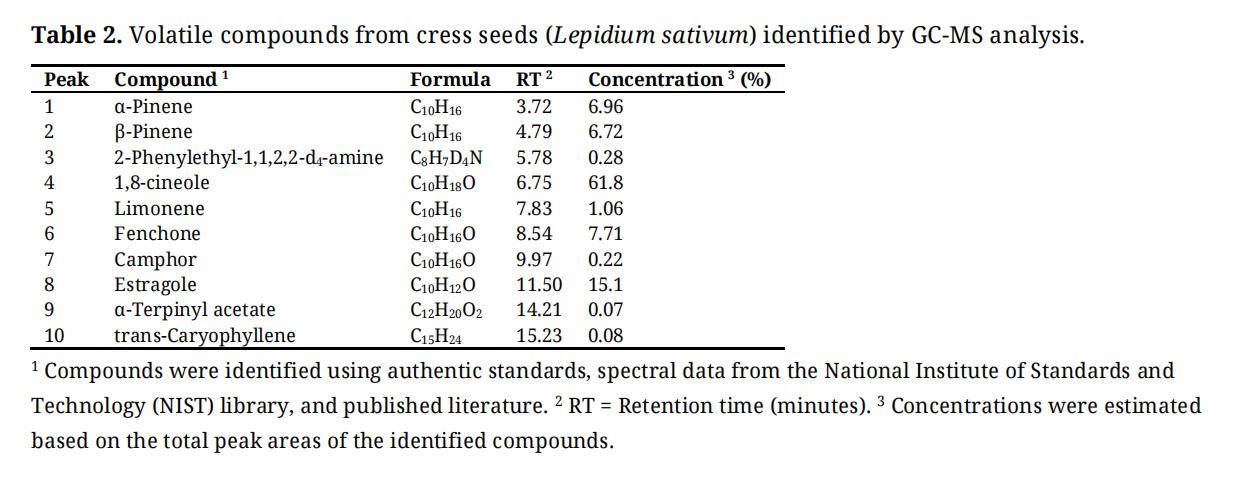 Table 2. Volatile compounds from cress seeds (Lepidium sativum) identified by GC-MS analysis.
Table 2. Volatile compounds from cress seeds (Lepidium sativum) identified by GC-MS analysis.
Figure 1 presents GP (mL/g) expressed per gram of DM, dDM, dNDF, and dADF. In vitro inclusion of cress seeds in the rumen fermentation substrate significantly influenced GP and greenhouse gas kinetics (Table 3). The asymptotic GP showed both linear (p < 0.001) and quadratic (p = 0.032) effects, with the quadratic trend being more pronounced, increasing by 50.2%, rising from 78.1 mL/g DM in the control to a peak value at 1.5% inclusion before a slight plateau at 2% cress seed inclusion. The rate of GP showed a significant linear decrease (p = 0.006) of approximately 9.2%, declining from 0.076 to 0.069 /h. Lag time before GP onset was not significantly affected (p = 0.705).
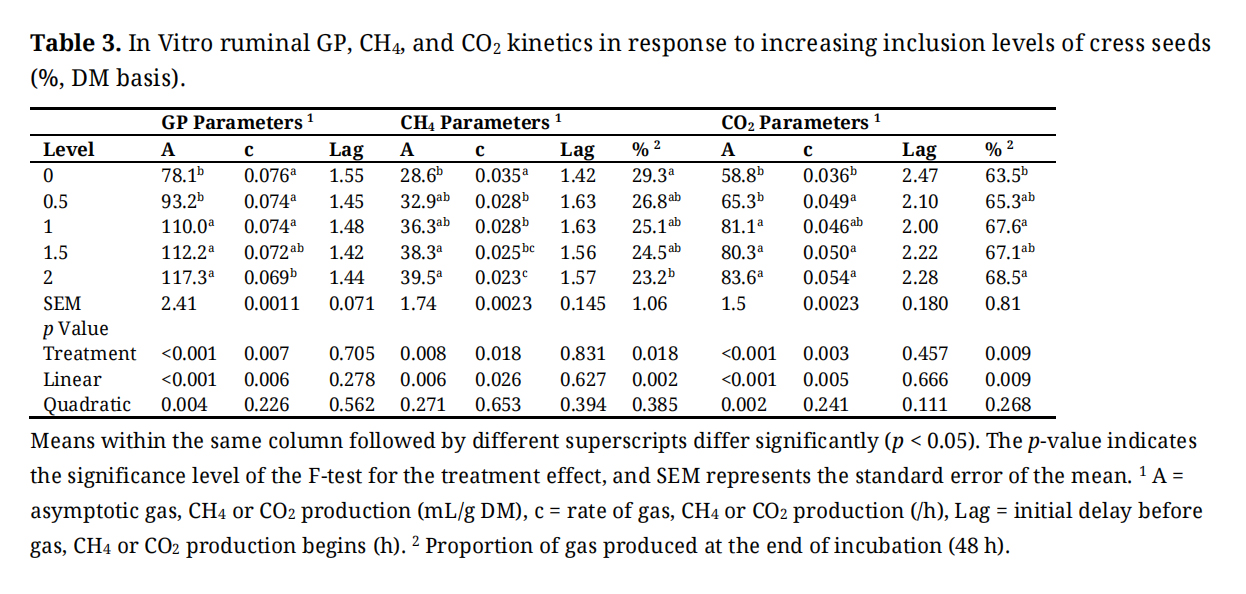 Table 3. In Vitro ruminal GP, CH4, and CO2 kinetics in response to increasing inclusion levels of cress seeds (%, DM basis).
Table 3. In Vitro ruminal GP, CH4, and CO2 kinetics in response to increasing inclusion levels of cress seeds (%, DM basis).
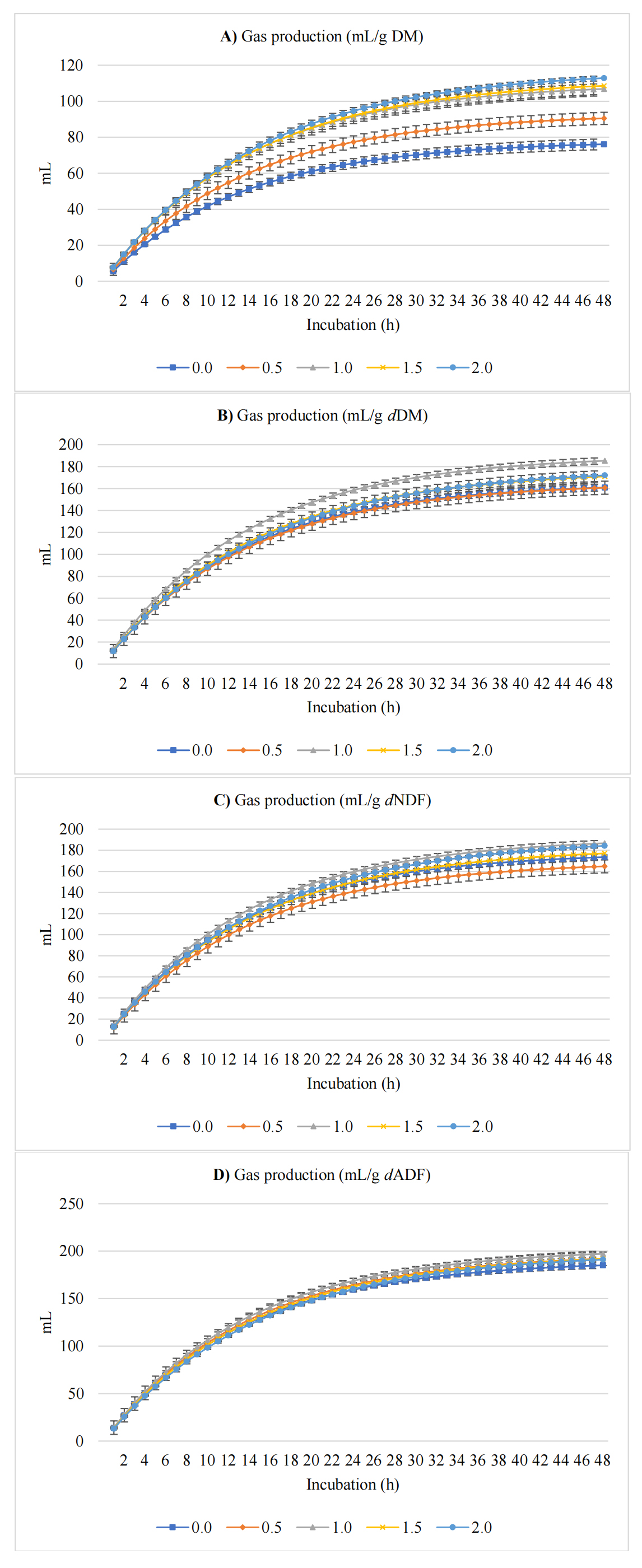 Figure 1. In vitro ruminal GP expressed as mL/g incubated DM (A), mL/g degradable DM (dDM) (B), mL/g degradable NDF (dNDF) (C), and mL/g degradable ADF (dADF) (D) of a TMR supplemented with varying levels of cress seeds.
Figure 1. In vitro ruminal GP expressed as mL/g incubated DM (A), mL/g degradable DM (dDM) (B), mL/g degradable NDF (dNDF) (C), and mL/g degradable ADF (dADF) (D) of a TMR supplemented with varying levels of cress seeds.
Figure 2 shows CH4 production (mL) expressed per gram of DM, dDM, dNDF, and dADF. Methane production also changed with cress seed levels. The asymptotic CH4 production followed a significant quadratic response (p = 0.021), increasing up to 1.5% inclusion and slightly declining thereafter, while the linear trend (p = 0.006) was less prominent. Overall, CH4 increased by 38.1%, from 28.6 mL/g DM in the control to 39.5 mL/g DM at 2% cress seeds inclusion. Conversely, the rate of CH4 production decreased linearly (p = 0.026) by 34.3% (from 0.035 to 0.023 /h), indicating a slower CH4 production rate despite higher total CH4 volume. The proportion of CH4 relative to total gas declined quadratically (p = 0.018), by 20.8%, from 29.3% in the control to a minimum at 1.5% inclusion before stabilizing at 23.2% at 2% cress seed inclusion. Lag time for CH4 production was unaffected (p = 0.831). The apparent shift in CH4 production observed at 12 h was due to the sampling intervals used during incubation; CH4 concentration was measured every 2 h up to 12 h, and then every 12 h (at 24, 36, and 48 h) thereafter.
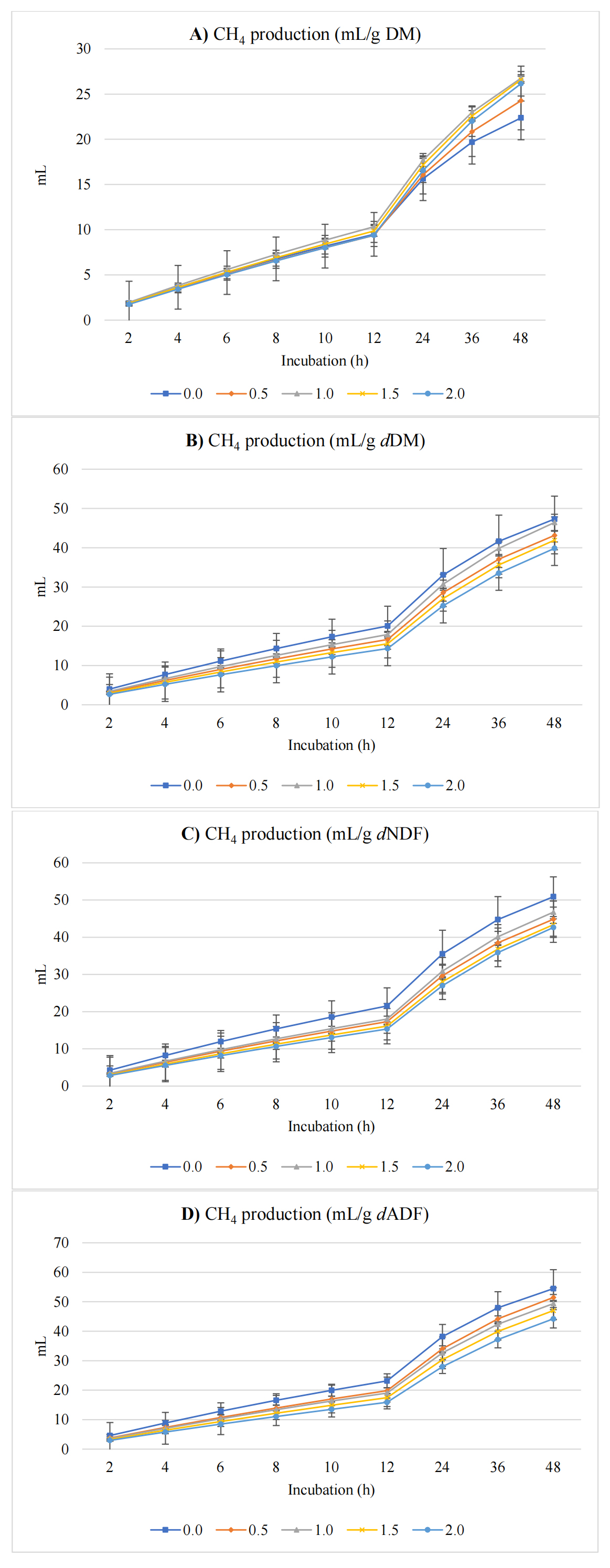 Figure 2. In vitro ruminal CH4 production expressed as mL/g incubated DM (A), mL/g degradable DM (dDM) (B), mL/g degradable NDF (dNDF) (C), and mL/g degradable ADF (dADF) (D) of a TMR supplemented with varying levels of cress seeds.
Figure 2. In vitro ruminal CH4 production expressed as mL/g incubated DM (A), mL/g degradable DM (dDM) (B), mL/g degradable NDF (dNDF) (C), and mL/g degradable ADF (dADF) (D) of a TMR supplemented with varying levels of cress seeds.
Figure 3 shows CO2 production (mL) expressed per gram of DM, dDM, dNDF, and dADF. CO2 production parameters increased significantly. The asymptotic CO2 production rose linearly (p < 0.001) and quadratically (p = 0.002) by 42.2%, from 58.8 mL/g DM in the control to a near-maximum at 1.5% and a slight plateau at 2% cress seeds inclusion. The rate of CO2 production also increased linearly (p = 0.005) by 50.0% (from 0.036 to 0.054 /h). The CO2 proportion relative to total gas increased linearly (p = 0.009) by 7.9% (63.5% to 68.5%). Lag time for CO2 was not significantly changed (p = 0.457).
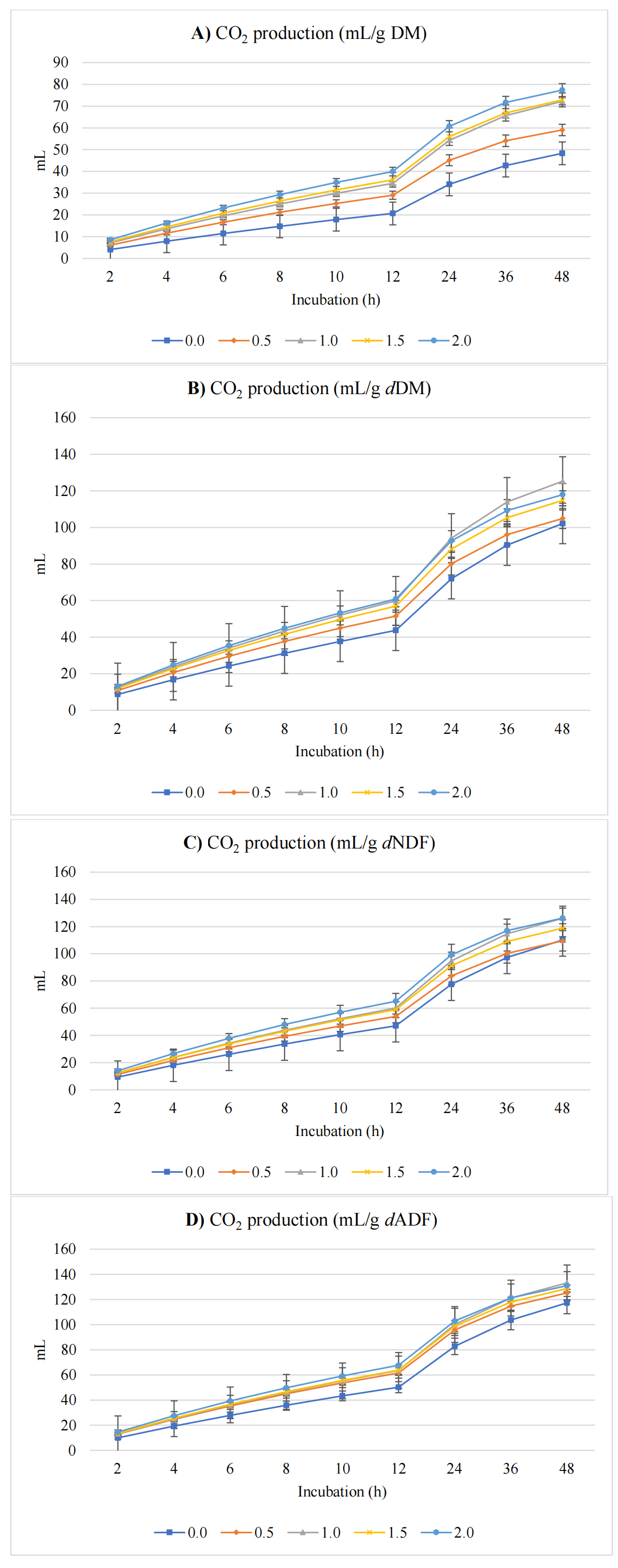 Figure 3. In vitro ruminal CO2 production expressed as mL/g incubated DM (A), mL/g degradable DM (dDM) (B), mL/g degradable NDF (dNDF) (C), and mL/g degradable ADF (dADF) (D) of a TMR supplemented with varying levels of cress seeds.
Figure 3. In vitro ruminal CO2 production expressed as mL/g incubated DM (A), mL/g degradable DM (dDM) (B), mL/g degradable NDF (dNDF) (C), and mL/g degradable ADF (dADF) (D) of a TMR supplemented with varying levels of cress seeds.
Supplementation of cress seeds significantly improved in vitro nutrient degradability and ruminal fermentation characteristics (Table 4). DM degradability increased markedly by 38.7% (quadratic response; p = 0.041), from 473 mg/g in the control to a peak of 656 mg/g at 1.5% inclusion before stabilizing at 2% cress seeds inclusion. Similarly, dNDF and dADF both exhibited significant quadratic responses (p < 0.05), rising from 439 to 614 mg/g and from 412 to 592 mg/g, respectively, indicating diminishing improvements beyond 1.5% inclusion.
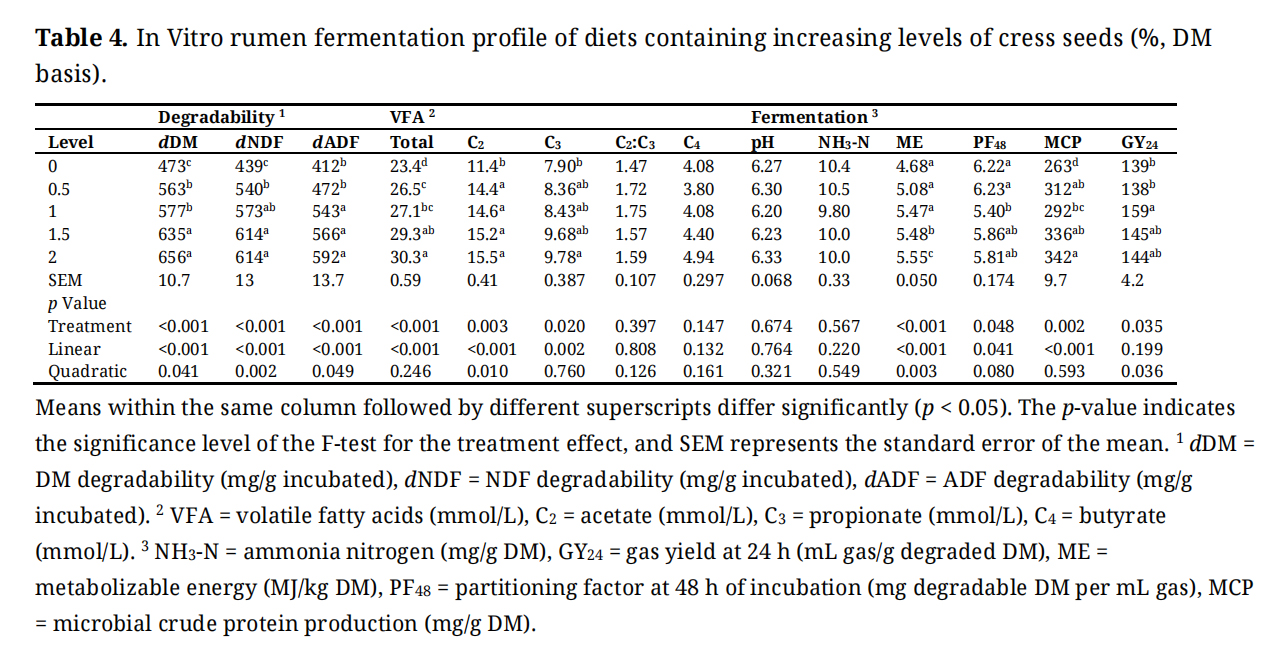 Table 4. In Vitro rumen fermentation profile of diets containing increasing levels of cress seeds (%, DM basis).
Table 4. In Vitro rumen fermentation profile of diets containing increasing levels of cress seeds (%, DM basis).
Total VFA concentration increased by 29.5% (p < 0.001; quadratic), from 23.4 to 30.3 mmol/L, with significant increases in acetate (C2) by 36.0% (p < 0.001; quadratic) and C3 by 23.9% (p = 0.002; linear). The C2:C3 ratio remained statistically unchanged (p = 0.397). Butyrate (C4) showed no significant effect (p = 0.147).
No significant differences were observed in ruminal pH (p = 0.674) or NH3-N concentrations (p = 0.567). However, ME increased significantly in a quadratic manner (p = 0.003), rising by 18.6% from 4.68 to 5.55 MJ/kg DM. Increasing levels of the additive significantly influenced both PF48 (p = 0.041; linear) and MCP production (p < 0.001; linear). The PF48 showed a significant quadratic decline (p = 0.041), with a 13.2% reduction at the 1% level compared to the control, followed by partial recovery at higher inclusion levels. Conversely, MCP increased linearly (p < 0.001), with enhancements of 18.6%, 27.8%, and a plateau beyond 1.5% inclusion. Additionally, GY24 was significantly affected in a quadratic pattern (p = 0.036), increasing from 139 to a peak of 144 mL/g DM at 1.5%, although the linear trend for this parameter was not significant.
The chemical composition of Lepidium sativum seed volatiles indicates a promising role as a natural feed additive for modulating rumen fermentation. The GC–MS analysis of Lepidium sativum (garden cress) seeds revealed a complex profile of bioactive volatile compounds, predominantly composed of oxygenated monoterpenes and phenylpropanoids. The most prominent compound identified was 1,8-cineole, accounting for 61.8% of the total volatile fraction. This high proportion is noteworthy, as 1,8-cineole (also known as eucalyptol) is a well-documented compound with potent antimicrobial, anti-inflammatory, and antioxidant properties [15]. In the context of ruminant nutrition, such compounds have been shown to modulate ruminal fermentation by selectively inhibiting methanogenic archaea and certain proteolytic or hyper-ammonia-producing bacteria [3,16], potentially contributing to a reduction in CH4 emissions and improved nitrogen utilization.
The substantial levels of estragole (15.1%), a phenylpropanoid known for its antimicrobial properties [17], also suggest a possible synergistic effect with 1,8-cineole in suppressing undesirable rumen microbes. Fenchone, α-pinene, and β-pinene, representing 7.71%, 6.96%, and 6.72% of the volatile content respectively, are monoterpenes frequently associated with modulation of rumen fermentation, especially by improving the C2:C3 ratio and enhancing fiber digestibility [18]. However, in the present study, the C2:C3 ratio was not significantly affected, possibly because the inclusion levels of cress seed were relatively low, leading to insufficient concentrations of active compounds to induce detectable shifts in fermentation pathways. Additionally, the microbial community may have adapted to the bioactive components without major alteration in acetate or propionate production. These compounds may exert selective pressure on cellulolytic and saccharolytic microbial populations, thereby affecting VFA profiles and potentially enhancing feed efficiency [3,16].
Although present at lower concentrations, minor compounds such as limonene, camphor, and α-terpinyl acetate contribute to the overall bioactivity of the essential oil matrix. Limonene has been reported to inhibit protozoal activity and reduce CH4 production [19], while camphor exhibits bacteriostatic effects on gram-positive organisms [20], which may translate into selective inhibition of some rumen microbiota. Additionally, the detection of trans-caryophyllene, a sesquiterpene with known anti-inflammatory and immune-modulating effects [21], highlights the potential systemic benefits of Lepidium sativum seed volatiles beyond the rumen environment. These effects align with the current interest in phytogenic feed additives as alternatives to synthetic rumen modifiers, such as monensin or ionophores, especially under systems aiming for antibiotic-free or organic livestock production [4].
In Vitro Gas, CH4, and CO2 Production KineticsA quadratic increase in asymptotic total GP and CO2 output with rising cress seed inclusion indicates an enhancement in the fermentative capacity of the rumen microbial ecosystem up to 1.5% inclusion, beyond which responses plateaued. Specifically, the 50.2% increase in GP and the 42.2% rise in CO2 volume at 2% inclusion reflect intensified microbial fermentation, likely driven by improved substrate degradability or a shift toward more efficient microbial pathways [22]. The significant quadratic responses in gas parameters indicate that cress seeds act in a dose-dependent manner, where beneficial effects are maximized before slight suppression occurs at excessive inclusion rates. Interestingly, despite the greater total gas output, the rate constant of GP decreased slightly (−9.2%), suggesting that while fermentation was more extensive, it proceeded at a slower pace. This could be attributed to the presence of bioactive compounds such as terpenes in cress seeds, which may exert selective antimicrobial effects. Specifically, these compounds can inhibit rapidly fermenting microbes, thereby allowing slower-growing, fiber-degrading populations to dominate the fermentation process [23].
Recently, Kholif et al. [18] reported similar findings with radish seeds, which are rich in 1,8-cineole (81.8%). Their inclusion led to an increase in both asymptotic GP and the rate of GP, indicating enhanced ruminal fermentation and improved dDM. Essential oils derived from plants—particularly those containing monoterpenoids like 1,8-cineole—have been shown to influence rumen fermentation and GP through their antimicrobial activity. The effectiveness of these phytochemicals is concentration-dependent and varies based on their specific chemical composition [24,25].
Higher asymptotic GP is typically associated with more effective utilization of fermentable substrates by rumen microbiota, leading to increased production of VFAs, which are vital energy sources for ruminants [26]. This enhanced fermentation contributes to better nutrient absorption, energy availability, and microbial protein synthesis—key factors supporting growth, milk production, and overall maintenance [27]. The trends observed in GP dynamics further suggest that the levels of cress seeds inclusion were within physiologically acceptable ranges, avoiding negative impacts on microbial activity or fermentation efficiency.
Moreover, certain volatile constituents of plant-derived essential oils, such as estragole and α-terpinyl acetate, may serve not only as antimicrobial agents but also as alternative carbon sources for specific rumen microbes, due to their hydrocarbon or alcohol-based structures [3]. The elevated concentrations of these volatile compounds in seeds like cress likely contribute to their modulatory effects on ruminal fermentation [28]. For example, eucalyptus oil—another essential oil rich in 1,8-cineole (eucalyptol)—has been shown to reduce total gas and CH4 emissions in vitro without impairing dDM and degradable OM, highlighting the dual role of these compounds in fermentation modulation and CH4 mitigation [29].
Furthermore, the observed rise in CO2 production can be explained by the intensified microbial degradation of fermentable carbohydrates. During ruminal fermentation, microbial metabolism of readily fermentable substrates produces CO2 alongside VFAs such as C2, C3, and C4 [30]. Therefore, the increase in total GP and CO2, coupled with decreased CH4 proportion, supports the hypothesis that the bioactive components of Lepidium sativum seeds modulate microbial activity in a selective manner. The significant rise in CO2 rate (50.0%) without a corresponding increase in lag time indicates that fermentative CO2 production was both quantitatively and kinetically stimulated, perhaps due to increased microbial decarboxylation or TCA cycle activity [31]. The higher CO2 proportion (from 63.5% to 68.5%) further reinforces the interpretation of an enhanced oxidative metabolic flux, which may be linked to a redirection of hydrogen away from methanogenesis [32].
The quadratic trend in CH4 production, with an increase up to 1.5% and a subsequent plateau, suggests that moderate levels of cress seed inclusion optimize fermentation efficiency while limiting excessive methanogenesis. This could be attributed to the bioactive compounds such as terpenes in cress seeds, which at moderate levels enhance microbial fermentation but at higher doses may partially inhibit some fermenters. Furthermore, the observed quadratic response of CH4 proportion, with a marked decline up to 1.5% inclusion, demonstrates a curvilinear improvement in fermentation efficiency, as less CH4 was produced per unit of gas, confirming more hydrogen utilization for alternative electron sinks such as propionate or microbial biomass [33,34].
This paradox—higher absolute CH4 emissions but reduced relative CH4 —may reflect a shift in fermentation dynamics due to the bioactive compounds in Lepidium sativum seeds. Terpenoids such as 1,8-cineole and estragole are known inhibitors of methanogenic archaea and protozoa [4], which likely redirected metabolic hydrogen toward alternative electron sinks such as microbial biomass or C3 synthesis [34]. Consequently, despite an increase in total fermentable substrate availability and total gas volume, a smaller fraction of the metabolic hydrogen was channeled into methanogenesis. The reduced rate of CH4 production supports this interpretation, suggesting a deceleration of hydrogenotrophic activity despite higher gas outputs.
A similar paradox has been observed with the inclusion of radish seeds, which are rich in 1,8-cineole [18]. While it was initially anticipated that these seeds would suppress methanogenesis by enhancing substrate degradation and optimizing rumen fermentation patterns, results indicated a linear increase in CH4 production with increasing inclusion levels. Methanogenesis in the rumen depends on a complex interplay between hydrogen-producing bacteria and methanogens, maintaining redox balance [34]. The enhanced CH4 output in such cases may be attributed to the high degradability of radish seeds, providing excess fermentable carbon sources and elevating microbial activity, including that of methanogens [4]. This was accompanied by a linear rise in total VFAs and C2 concentrations without a corresponding increase in C3 levels, which aligns with higher CH4 emissions, as C2 fermentation generates excess hydrogen subsequently utilized by methanogens [35,36].
These results are consistent with previous studies showing that monoterpene-rich essential oils (e.g., α-pinene, limonene) can suppress methanogens and hyper-ammonia-producing bacteria while enhancing fibrolytic microbes [3,16,37]. The improved GP profile and reduced CH4-to-gas ratio observed in this study support a beneficial shift in rumen microbial ecology, though confirmation via microbial community analysis or rumen metabolomics is warranted.
In Vitro Rumen Fermentation ProfileThe supplementation of cress seeds (Lepidium sativum) in a TMR significantly influenced key parameters of in vitro rumen fermentation, showing predominantly quadratic responses suggest enhanced nutrient utilization and microbial activity. The observed linear and quadratic improvements in dDM, dNDF, and dADF align with increased microbial colonization and fiber breakdown efficiency, as indicated by Dhiman et al. [38]. Specifically, the increase in dDM and the improvements in fiber degradability point to the beneficial effects of bioactive compounds in cress seeds—particularly oxygenated monoterpenes such as 1,8-cineole, estragole, and fenchone—on the rumen microbial ecosystem [4,37].
These findings are consistent with those of Kholif et al. [4], who reported similar improvements in asymptotic GP and fermentation kinetics with radish seed supplementation rich in 1,8-cineole. The results from the current study and that of Kholif et al. [4] suggest that monoterpenes can stimulate microbial growth and fibrolytic activity by modulating rumen microbial populations. Terpenoids and phenylpropanoids have been shown to selectively inhibit methanogenic archaea and protozoa, which typically suppress fibrolytic bacteria through predation [4,39]. The reduction in protozoal pressure may thus indirectly favor fiber-degrading bacteria such as Fibrobacter succinogenes and Ruminococcus spp., leading to greater fiber digestibility [39,40].
The quadratic nature of the response indicates that inclusion beyond 1.5% offers no additional benefit, possibly due to microbial saturation or mild inhibitory effects at high concentrations [41]. This emphasizes the importance of optimizing inclusion rates to maximize benefits without adversely affecting the microbial balance.
The quadratic increase in total VFAs by 29.5% reflects a significant improvement in carbohydrate fermentation and energy yield, a core driver of microbial growth and ruminant performance [42]. The specific rise in C2 and C3, without altering the C2:C3 ratio, suggests simultaneous enhancement of both structural and non-structural carbohydrate fermentation pathways [34]. This balanced fermentation supports de novo fatty acid synthesis in the mammary gland (via C2) and gluconeogenesis for energy supply (via C3), both crucial for high-producing ruminants [43].
In parallel, similar effects were reported with radish seeds, where total VFA concentrations increased with supplementation, signifying improved fermentability and microbial energy production [11]. Acetate production, in particular, increased in response to higher inclusion levels and is known to contribute 50–70% of total VFAs in the rumen, playing a critical role in milk fat synthesis and energy metabolism [34,43].
Importantly, MCP production quadratically increased significantly—up to 30% at 2% inclusion—demonstrating enhanced microbial protein synthesis efficiency. This improvement is linked to greater ATP yield from fermentable substrates and better nitrogen utilization, both of which are essential for maximizing the flow of high-quality protein to the host animal [44]. Similar improvements in MCP were reported with radish seed supplementation, where increased fermentable carbohydrates and a favorable microbial environment supported microbial biomass production [18]. Comparable results have also been documented in studies using essential oils or plant extracts rich in 1,8-cineole, such as Rosmarinus officinalis and Salvia officinalis, which enhanced microbial growth efficiency and nitrogen assimilation in the rumen [22,45]. These findings reinforce the concept that bioactive phytochemicals in seeds stimulate microbial populations involved in protein synthesis.
Interestingly, while PF48 declined linearly at 1% inclusion (13.2% decrease)—the partial recovery at higher levels suggests not a time-dependent adaptation, since all treatments were incubated for the same duration, but rather a microbial compositional adjustment or a shift in substrate–microbe interactions [22]. This mirrors previous observations where functional feed additives modulated fermentation efficiency through microbial community restructuring and selective enrichment of tolerant populations, rather than through temporal adaptation. The significant quadratic response in GY24, despite a non-significant linear trend, suggests a complex interaction between fermentable substrates and microbial dynamics, consistent with findings on radish seeds [18].
Ruminal pH and NH3-N levels remained stable, indicating that fermentation enhancement did not compromise ruminal buffering capacity or NH3-N dynamics [46]. The stable pH suggests effective VFA absorption and/or buffering [47], while unchanged NH3-N concentrations imply balanced proteolysis and nitrogen assimilation. Although previous studies with radish seeds reported reduced NH3-N due to 1,8-cineole-mediated suppression of hyper-ammonia-producing bacteria, such an effect was not observed here—potentially due to lower antimicrobial thresholds in cress or differences in fermentation substrates [3,16,48].
Nevertheless, the concurrent increase in MCP and stable NH3-N values suggests efficient NH3-N incorporation into microbial protein, indicating a highly synchronized energy-nitrogen balance in the rumen. This is essential for maximizing microbial growth and feed efficiency [11]. As observed with radish supplementation, the presence of bioactive phytochemicals likely contributed to improved microbial efficiency and nitrogen retention [22].
Additionally, the increased ME (+18.6%) and enhanced GY24 further support the improved fermentative efficiency of the rumen ecosystem. These effects are likely driven by the fermentable carbohydrate content and stimulatory effects of phytochemicals on microbial metabolism [49]. It is worth noting that while higher GP may raise concerns about CH4 emissions, the balanced SCFA profile and increased MCP observed here suggest a shift toward more efficient fermentation pathways rather than energy loss.
Overall, these quadratic responses confirm that cress seed inclusion exerts a dose-dependent effect, enhancing fermentation and microbial growth up to an optimal level (1–1.5%), after which responses stabilize or slightly decline. This pattern supports the reviewer’s interpretation that the highest significant polynomial trend—the quadratic effect—best explains the fermentation dynamics observed.
Finally, phytochemicals in cress may modulate microbial communities by suppressing non-beneficial organisms (e.g., methanogens and protozoa) while supporting beneficial fibrolytic and proteolytic bacteria. These microbial shifts contribute to improved fermentation, degradability, and energy capture, especially in high-fiber diets. Further in vivo studies, coupled with rumen microbiome analyses, are recommended to validate these results under practical feeding conditions and clarify the ecological interactions induced by cress bioactives.
Cress seed inclusion significantly enhanced fermentation dynamics. Total gas and CO2 production increased quadratically, while the proportion of CH4 in total gas declined by 20.8%, despite an increase in absolute CH4 volume. Methane production rate decreased by 34.3%, suggesting more efficient fermentation with delayed methanogenesis. In terms of nutrient utilization, supplementation linearly improved DM (+38.7%), NDF (+39.9%), and ADF (+43.6%) degradability, along with total VFAs (+29.5%), ME (+18.6%), and MCP synthesis (+4.3%). No adverse effects were observed on ruminal pH or NH3-N nitrogen levels, confirming the stability of the rumen environment. Based on these findings, dietary inclusion of cress seeds at 2% of diet DM is recommended to maximize fiber degradation, enhance microbial protein yield, and reduce CH4 intensity, making it a promising natural additive for improving ruminant productivity and sustainability.
All data generated from the study are available in the manuscript.
Conceptualization, AEK, UYA and TAM; Data curation, AEK, TAM and GAG; Formal analysis, AEK and TAM; Funding acquisition, AEK; Investigation, AEK, TAM and GAG; Methodology, AEK, TAM and GAG; Project administration, AEK; Resources, AEK and GAG; Software, AEK, MJA and TAM; Supervision, AEK and TAM; Validation, AEK, UYA and GAG; Visualization, AEK and TAM; Writing—original draft, AEK and UYA; Writing—review and editing, AEK and UYA. All authors have read and agreed to the published version of the manuscript.
The authors declare that there are no conflicts of interest.
This research received no external funding.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
Kholif AE, Gouda GA, Morsy TA, Anele UY. Sustainability of using cress seed as a feed additive: Effects on in vitro gas production, methane emissions, and rumen fermentation. J Sustain Res. 2025;7(4):e250064. https://doi.org/10.20900/jsr20250064.

Copyright © Hapres Co., Ltd. Privacy Policy | Terms and Conditions




